

当前位置:易推广 > 上海岑特生物科技有限公司>公司新闻>你知道喂给细胞的培养基成分去哪了吗?
企业档案
会员类型:会员

已获得易推广信誉 等级评定
(0 -40)基础信誉积累,可浏览访问
(41-90)良好信誉积累,可接洽商谈
(91+ )优质信誉积累,可持续信赖
易推广会员:8年
工商认证 【已认证】

最后认证时间:
注册号:91310120MA1HL6**** 【已认证】
法人代表: 侯亚*** 【已认证】
企业类型:经销商 【已认证】
注册资金:人民币**万 【已认证】
产品数:20155
参观次数:6249109
手机网站:http://m.yituig.com/c128110/
旗舰版地址:http://guduo.app17.com
公司新闻
你知道喂给细胞的培养基成分去哪了吗?
点击次数:811发布时间:2019/12/16
我们做科研的,大多数人都养过细胞,细胞样本实验周期短,成本低,我们在养细胞的时候不可避免的要用到培养基,比如微生物用到的 LB, M9 等,动物细胞用到的 DMEM 等,每种培养基的成分都不尽相同,那么你有没有考虑过,培养基是如何给细胞提供营养的呢?那些 C 源 N 源到底是怎么维持细胞生长的呢?我们今天要分享的这篇文献,就以谷氨酰胺为例,使用同位素追踪的方法给大家展示在缺氧条件下,谷氨酰胺作为 C 源是如何维持肿瘤细胞生长的。
部分 介绍谷氨酰胺的重要性
当然了,先证明谷氨酰胺对细胞生长是很重要的,再来研究他的代谢,没毛病。
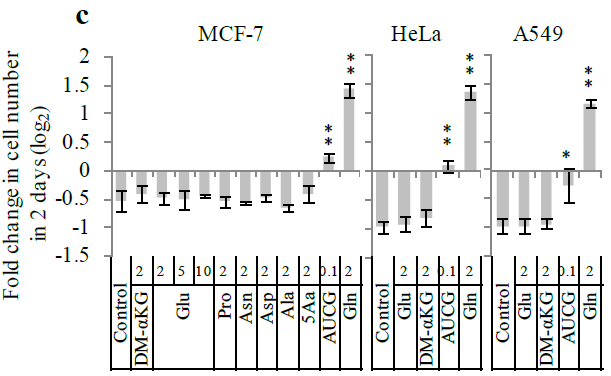
首先用添加了不同浓度、不同成分的培养基(Glu,AUCG,各种氨基酸)培养肿瘤细胞,发现培养基中若没有谷氨酰胺,细胞死亡率会显著上升,而添加了 AUCG 的培养基可以在谷氨酰胺缺失的情况下抑制细胞的死亡,说明谷氨酰胺与核苷的合成有关。由此,作者推论,谷氨酰胺是细胞生长所必须的物质。
那么在缺氧条件下细胞会将谷氨酰胺作为 C 源维持生长,但是谷氨酰胺也是生物体的 N 源,那么细胞会如何处理随之而来的高浓度的 N 呢?
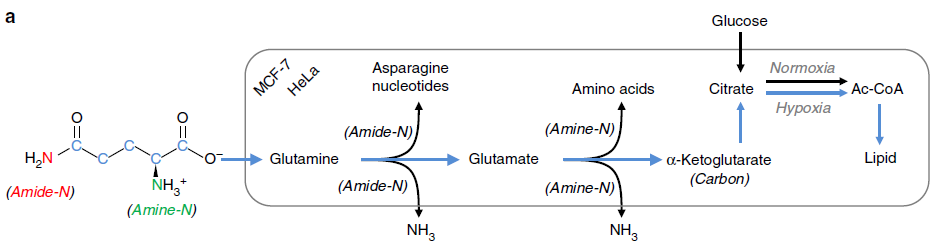
在缺氧条件下谷氨酰胺被用来做为主要的 C 源,尤其是脂质合成。
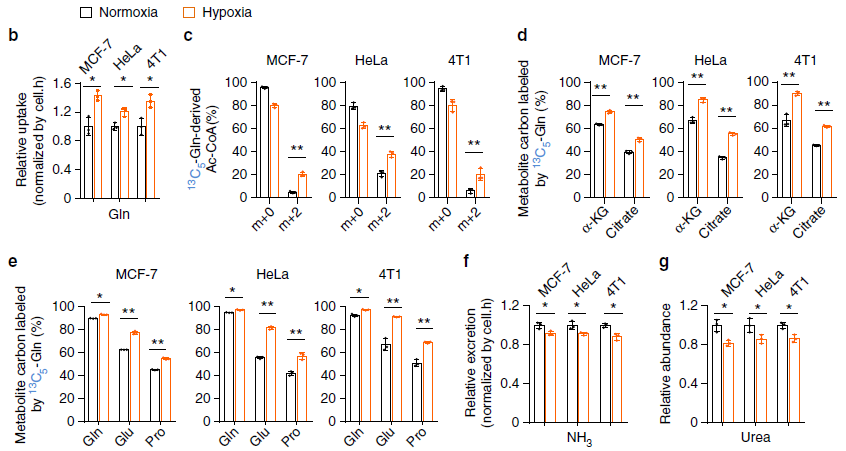
由上图 b 可知缺氧条件下细胞对谷氨酰胺(Gln)吸收增加了。通过 13C-Gln 标记发现脂质前体 Ac-CoA 增加了(图 c,m+0 减少,m+2 增加),其他下游物质如氨基酸等也增加了(图 d,e)。这些数据表明缺氧时谷氨酰胺作为 C 源吸收增加了。接下来,作者试图找到谷氨酰胺的 N 代谢。谷氨酰胺可以脱去氨基生成 NH3,但是对培养基的检测发现 NH3 的量没有升高反而下降了(图 f),NH3 可以被代谢成尿素,但是培养基中尿素的含量却下降了(图 g),因此谷氨酰胺的 N 应该是被其他物质利用了。
第二部分 研究谷氨酰胺的代谢
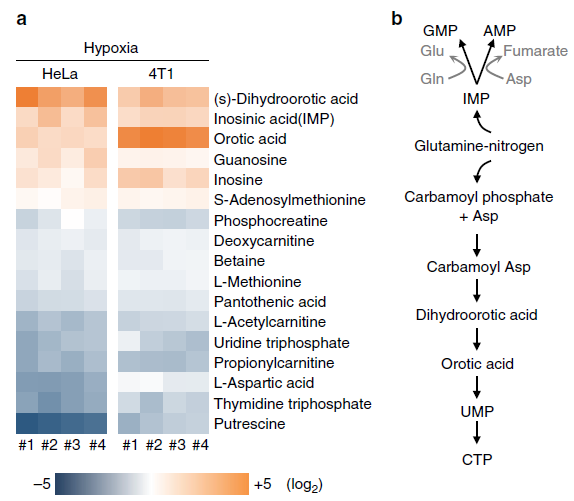
通过在缺氧条件下的含 N 代谢物的筛选,得到了 6 个显著升高的物质,11 个显著下降的物质。其中,五个显著上调的物质:IMP、DOHA、OA、鸟嘌呤和肌苷参与了 nucleotide biosynthesis 通路,验证结果也表明在 HeLa、MCF-7 和 4T1 细胞中 IMP(AMP 和 GMP 的前体)、carbamoyl-Asp、DHOA、OA 都显著升高。这些结果表明缺氧导致细胞内核苷酸的前提累积,尤其是嘧啶前体,包括 carbamoyl-Asp、DHOA 和 OA。
・缺氧条件下谷氨酰胺的 N 富集在 DOHA 和 OA 上
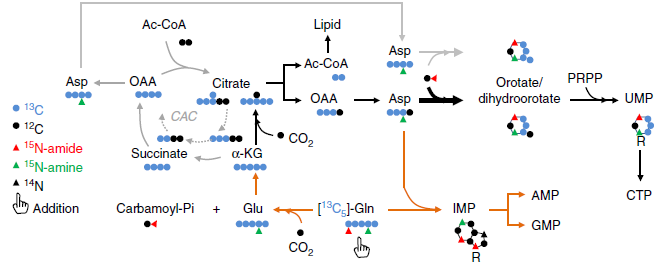
作者使用 N15 标记的谷氨酰胺来追踪 N 的吸收。由上图可知,酰胺基(Amine)上的 N 会转移到 Asp、DOHA、OA、UMP、IMP 上,而氨基(Amide)上的 N 会转移到 DHOA、OA、UMP、IMP 上。
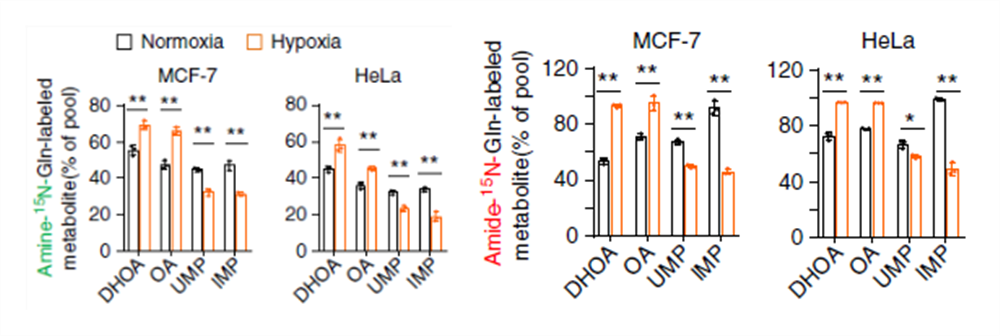
在缺氧条件下,标记的 DHOA 和 OA 含量显著上升,说明谷氨酰胺的 N 富集于 DHOA 和 OA 上,但是 DHOA 和 OA 的代谢效率不高,因为下游产物 UMP 含量没有增加。
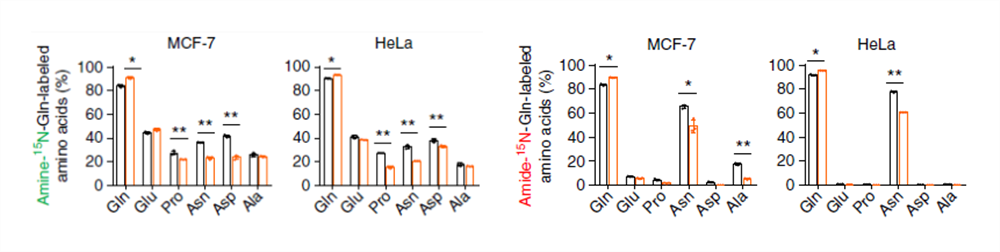
标记的 N 还可以在氨基酸中检测到,但是在缺氧条件下,被标记的氨基酸含量都没有升高,因此这些数据可以证明缺氧条件下谷氨酰胺的 N 富集于 DHOA 和 OA 上而不是氨基酸上。
・缺氧条件下促使天冬氨酸作为嘧啶前体
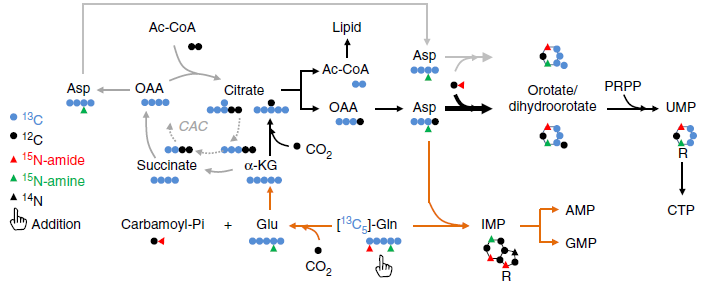
由上图可知,谷氨酰胺的 C 在通过 TCA 循环和 α- 酮戊二酸羧化还原途径转移到天冬氨酸之后可以被合成为嘧啶。这两条途径可以通过检测 C13-Gln 来源的乙酰辅酶 A,天冬氨酸,柠檬酸,DHOA 和 OA 来区分。正常条件下,谷氨酰胺的 C 进入 TCA 循环,可以生成 M+4 的 Asp,柠檬酸,DHOA 和 OA;在缺氧条件下,生成 m+2 的乙酰辅酶 A,m+3 的天冬氨酸,m+5 的柠檬酸,m+3 的 DHOA 和 OA 通过还原途径生成;m+3 的 UMP 可以由两条途径都生成。
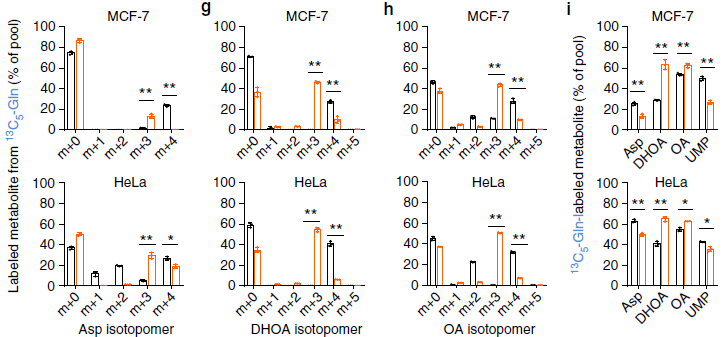
由上图结果可知缺氧条件下谷氨酰胺的 C 通过还原途径进入了乙酰辅酶 A、DHOA 和 OA,并且含量有所升高,因此缺氧条件促进了他们的合成。但 DHOA 的直接前体:天冬氨酸含量却降低了,这可能与嘧啶合成通路中多个酶参与有关,为研究这个现象,作者使用 C13,15N-Asp 来追踪天冬氨酸在嘧啶合成中的代谢。
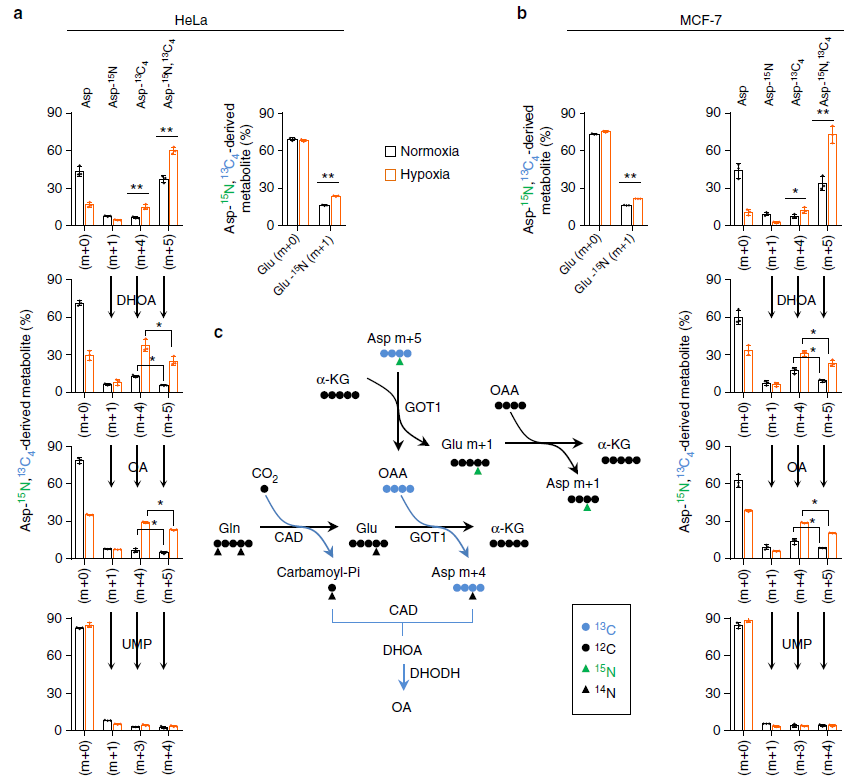
由上图可知,检测到了 m+5 的 Asp 表明细胞完全可以吸收外源的 13C-15N-Asp(图 a),也检测到了 15N-m+1 的谷氨酸和天冬氨酸,13C-m+4 的天冬氨酸(图 a,b);缺氧使细胞吸收更多的天冬氨酸,有趣的是,正常条件下细胞并不能产生很多的 DHOA 和 OA, 但是在缺氧条件下 DHOA 和 OA 的含量都升高了(m+4,m+5 显著升高),而且 m+4 的天冬氨酸生成 m+5 的 DHOA 和 OA 效率更高了,因此缺氧使天冬氨酸到 DHOA 和 OA 的途径活性升高了,这也导致细胞内的天冬氨酸含量降低。
・DHOA 和 OA 是细胞生长必须的吗?
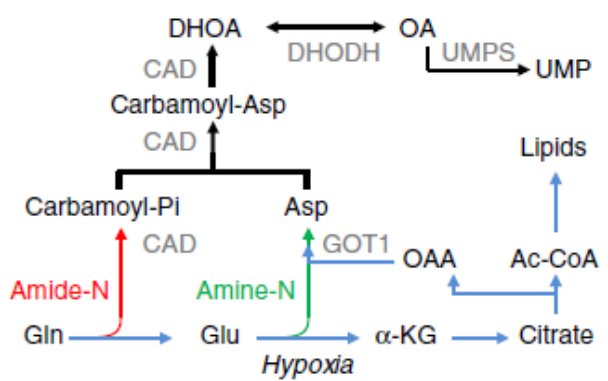
从上述结果来看,缺氧条件下大部分的谷氨酰胺都生成了 DHOA 和 OA,那么他们对细胞生存是必须的吗?作者对 DHOA 和 OA 通路上的酶做了 knockdown,如 CAD、DHODH、GOT1。
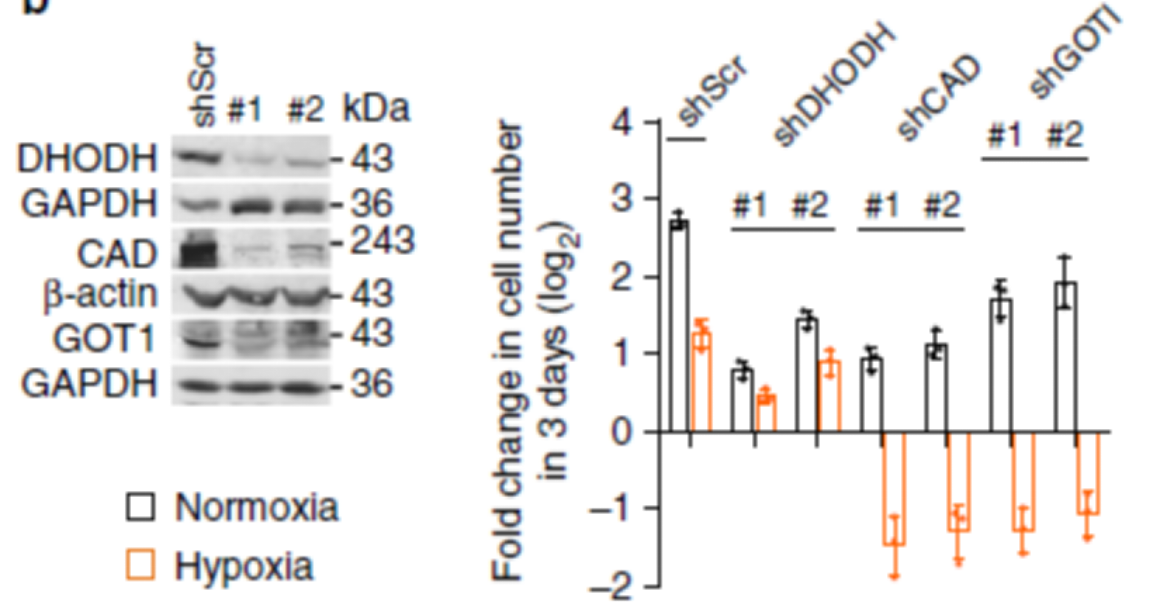
DHODH 和 CAD 是嘧啶合成的关键酶,因此他们的 knockdown 即使在正常条件下也明显的影响了细胞的分化。相反的是,缺氧条件下,GOT1 的敲除轻微地影响了细胞的分化。而 CAD 和 GOT1 的敲除让细胞无法生长,DHODH 的敲除只是稍微影响细胞的生长。这说明对于细胞来说,DHOA 才是不可缺少的,而不是 OA。
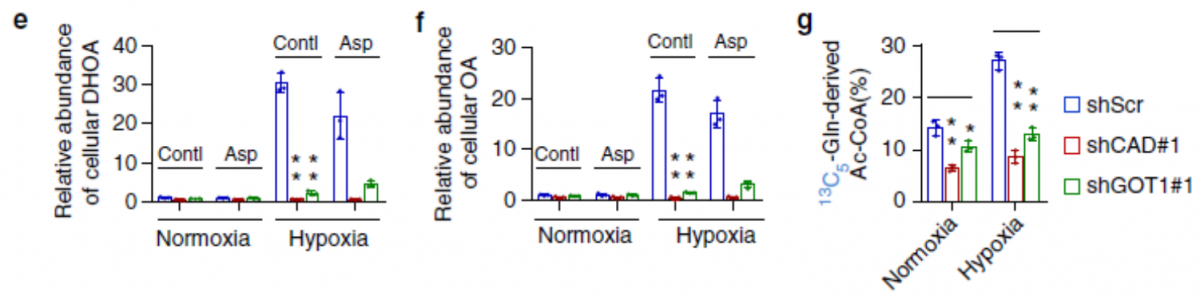
CAD 和 GOT1 的敲除显著降低了 DHOA 和 OA 的含量(图 e,f),同时也减少了 α- 酮戊二酸,柠檬酸和乙酰辅酶 A 的含量(图 g)。虽然天冬氨酸可以合成 DHOA 和 OA,外源添加天冬氨酸也并没有使 DHOA 和 OA 的量增加(图 e,f)。这些数据说明谷氨酰胺的 C 代谢和 N 代谢都与 DHOA 有关。
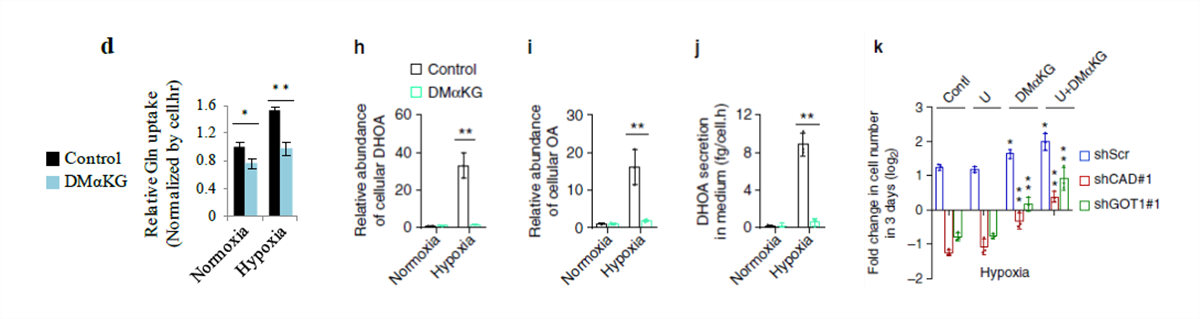
为了确认这个假设,使用 α- 酮戊二酸(C 来自谷氨酰胺)处理细胞,结果显示 α- 酮戊二酸的添加可以减少谷氨酰胺的吸收(图 d),几乎完全清除了 DHOA 和 OA 的积累(图 h,i),并且抑制了 DHOA 的分泌(图 j)。尿苷和 α- 酮戊二酸的添加可以使敲除酶的细胞增值增加,但是单独添加尿苷不能,单独添加 α- 酮戊二酸或添加 α- 酮戊二酸和尿苷可以升高细胞的增殖,这些结果表明谷氨酰胺的 C 代谢对细胞生长来说是必要的,并且取决于 DHOA 的合成。
总结一下
通过同位素追踪,文章展示了一条在缺氧条件下的谷氨酰胺通路,找到了缺氧条件下肿瘤细胞中的代谢变化,找到了谷氨酰胺的代谢产物 DHOA 和 OA, 并在病人和小鼠中也检测到了升高的 DHOA 和 OA,因此他们可以作为肿瘤的潜在标志物,通路上的酶如 CAD 和 GOT1 可以作为肿瘤治疗的可能目标。
怎么样,代谢流的结果是不是很强大呢?
它可以为我们提供更直接的证据来说明问题,在基因工程和不同环境或不同代谢途径中找到关键通路及代谢功能变化方面提供强有力的证据。
相关产品
-
 电议 型 号:
电议 型 号: -
电议 型 号:
-
电议 型 号:
-
电议 型 号:
-
电议 型 号:


