
如同爱情是文学永恒的话题,长生不老也是人类永恒的话题。 近年来,与抗衰老相关的各类“神药”甚嚣尘上,如李嘉诚投资的烟酰胺核糖 (Nicotinamide Riboside; NR)。NR 是烟酰胺腺嘌呤二核苷酸 (NAD+) 前体,可以提高人体内 NAD+ 水平。众所周知,NAD+ 在衰老过程中会下降,这是为什么呢?上个月 Nature Metabolism 同期两篇研究揭示了其中的一项重要原因:衰老诱导的炎症会促进 CD38 在免疫细胞中的积累或激活,增加了 NAD+ 消耗,从而引起 NAD+ 水平下降。 其实,衰老过程与许多其他生物学过程一样,受到经典信号通路和转录因子的调控。事实上,科学家们通过对不同生物系统和信号通路的干预,已经在多种动物模型中改变衰老的进程,延缓与衰老相关的多种疾病的发生。 目前,已经确定了几种衰老相关的关键信号通路,包括 Insulin/IGF-1 signaling (IIS)、mTOR、AMPK、NF-κB 和 Sirtuins 通路。这些信号传导途径感应营养物质或代谢产物,调节葡萄糖、氨基酸、cAMP 和 NAD+ 的水平,并形成复杂的与长寿和衰老相关的网络。
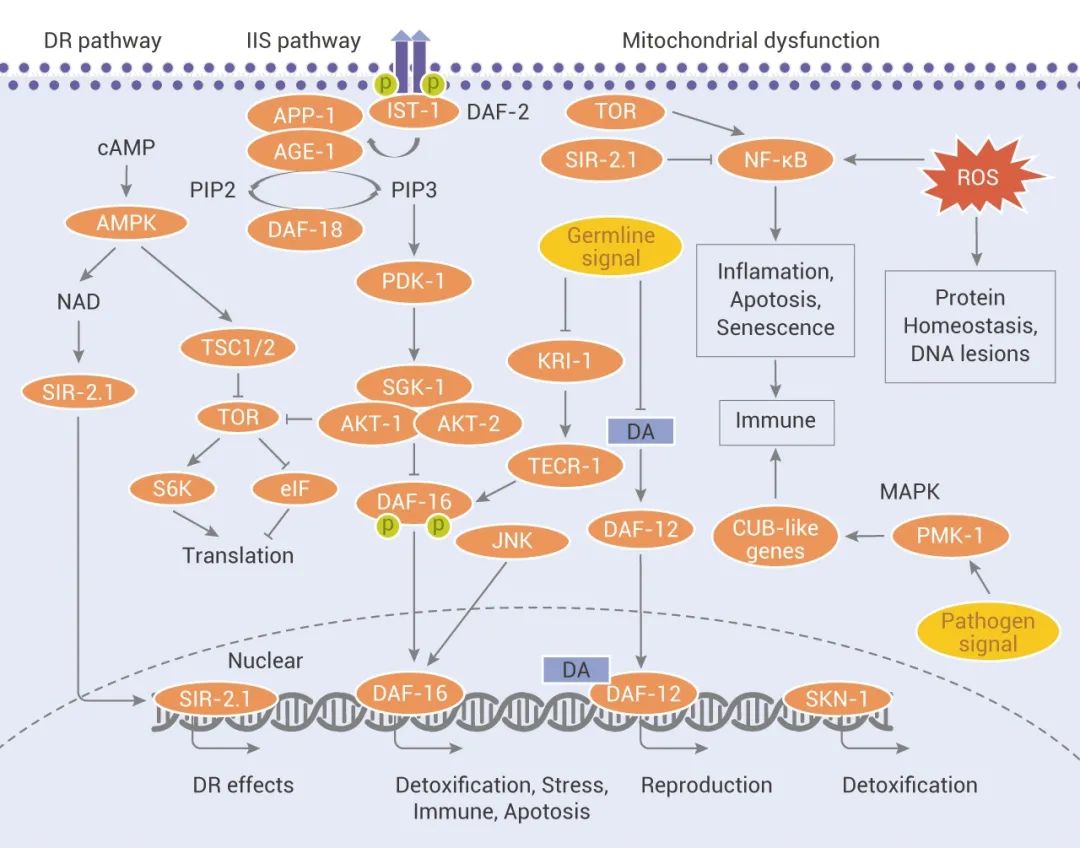
图 1. 衰老相关的信号网络[4]
■ Insulin/IGF-1 signaling (IIS) 信号传导
IIS 信号传导途径是在模型生物中*早定义的与衰老和年龄有关的途径。胰岛素/胰岛素样肽 (ILP) 与目标细胞表面表达的胰岛素受体结合,从而触发经由 IIS 途径的信号转导。这将启动细胞内激酶级联反应,*终导致激酶 AKT 的激活。AKT 激活后则磷酸化下游转录因子 FOXO,这会抑制 FOXO 的转录功能,从而促进细胞存活、生长和增殖。另外,IIS 信号通路还与 mTOR、AMPK 等通路相互关联,形成一个复杂的调控寿命和衰老的网络。 越来越多的证据表明,生长激素/胰岛素样生长因子-1 (GH/IGF-1) 信号通路也在调节衰老和疾病中起重要作用。在哺乳动物中,生长激素可诱导肝脏中 IGF-1 的释放,IGF-1 通过与细胞表面的高亲和性 IGF-1R 结合,并激活胰岛素受体底物分子 (IRS) 磷酸化和 PI3K-Akt 途径及 MAPK 信号级联,来启动复杂的细胞内信号级联,控制多个功能包括 mTOR 活性和 FOXO 易位。 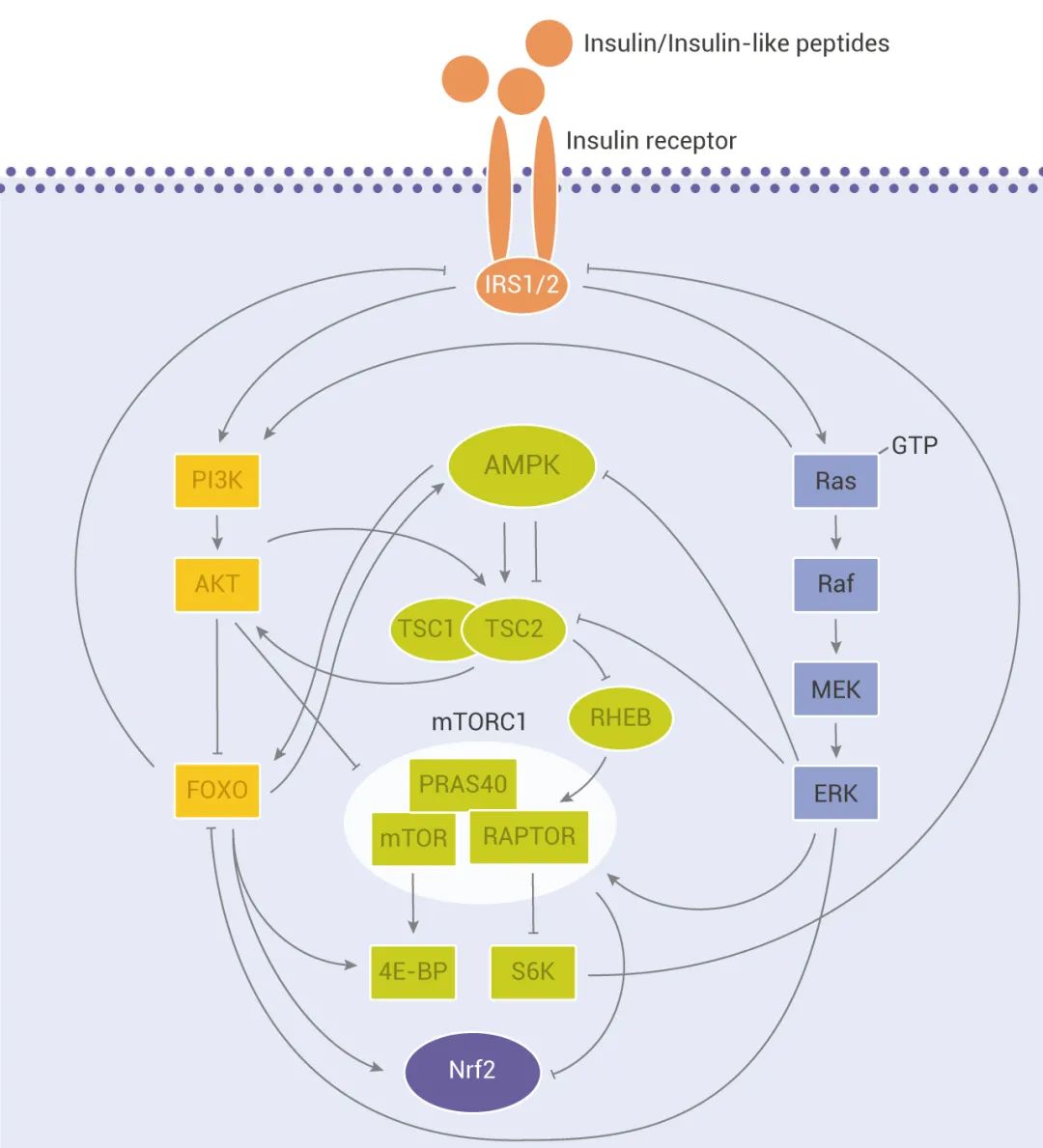
图 2. IIS 信号通路与其他长寿相关的细胞信号网络[3]
mTOR 是一种进化保守的丝氨酸-苏氨酸激酶,可感知并整合多种环境和细胞内信号,例如生长因子和营养物质,以控制蛋白质和脂质合成或自噬等过程。越来越多的证据表明,mTOR 信号会影响寿命和衰老。
mTORC1 调控的衰老相关分泌表型 (SASP)。PS:衰老细胞分泌促炎介质以促进衰老,被称为 SASP。mTORC1 的抑制剂 Rapamycin 可降低 IL-6 和其他细胞因子 mRNA 水平,并选择性抑制膜结合的细胞因子 IL-1A 的翻译。IL-1A 的减少降低了 NF-κB 的转录活性,从而控制了大部分的 SASP。 此外,mTOR 还通过控制着 MAPKAPK2 (MK2) 的翻译来调控 SASP。这是由于在衰老过程中 MAPKAPK2 使 RNA 结合蛋白 ZFP36L1 磷酸化,从而抑制其降解众多 SASP 组分的转录本的能力。此外,在衰老小鼠和细胞中,两种重要的 DNA 损伤修复蛋白 MGMT 和 NDRG1 受到 mTORC1 的负调控。 值得注意的是,IIS 通路可与 mTOR 相互联系调节衰老,例如 IIS 通过 AKT 激活了 mTORC1,而 mTORC1 可以通过 S6K 负调控 IIS,从而抑制了胰岛素受体底物 1 (IRS-1)。研究表明,降低 IIS 信号转导可延长线虫、果蝇和小鼠的寿命。
AMPK 在细胞和有机体的能量代谢中起基础性作用。由能量不足引起的 ATP 消耗激活 AMPK 信号,而 AMPK 信号反过来又刺激分解代谢过程来维持能量稳态,相应的,AMPK 关闭了许多与能量消耗有关的反应,如蛋白质和脂类合成。研究表明,AMPK 信号的激活能力随着年龄的增长而下降,这会损害细胞内环境的有效平衡,并加速衰老过程。因此,AMPK 信号的激活受损会干扰下游信号网络的功能,从而导致细胞内稳态维持的问题。
研究表明 AMPK 信号通过作用于下游,控制着复杂的信号网络,包括几个长寿相关通路,如 FOXO、mTOR/ULK1 和 SIRT1 信号。大量文献表明,IIS 信号通路抑制 AMPK 的激活,进而降低其下游活性。 另外,AMPK 可激活 ULK1,抑制增强自噬的 mTOR 信号。而随着年龄的增长,AMPK 信号的下降会干扰自噬和细胞蛋白质稳态的维持。鉴于 AMPK 在能量代谢中具有如此重要的作用,与年龄相关的线粒体内稳态恶化可能也是由 AMPK 活化效率低下引起的。此外,AMPK 信号可以抑制 NF-κB 系统,从而控制炎症和免疫反应,后者随着年龄的增长而受到干扰。
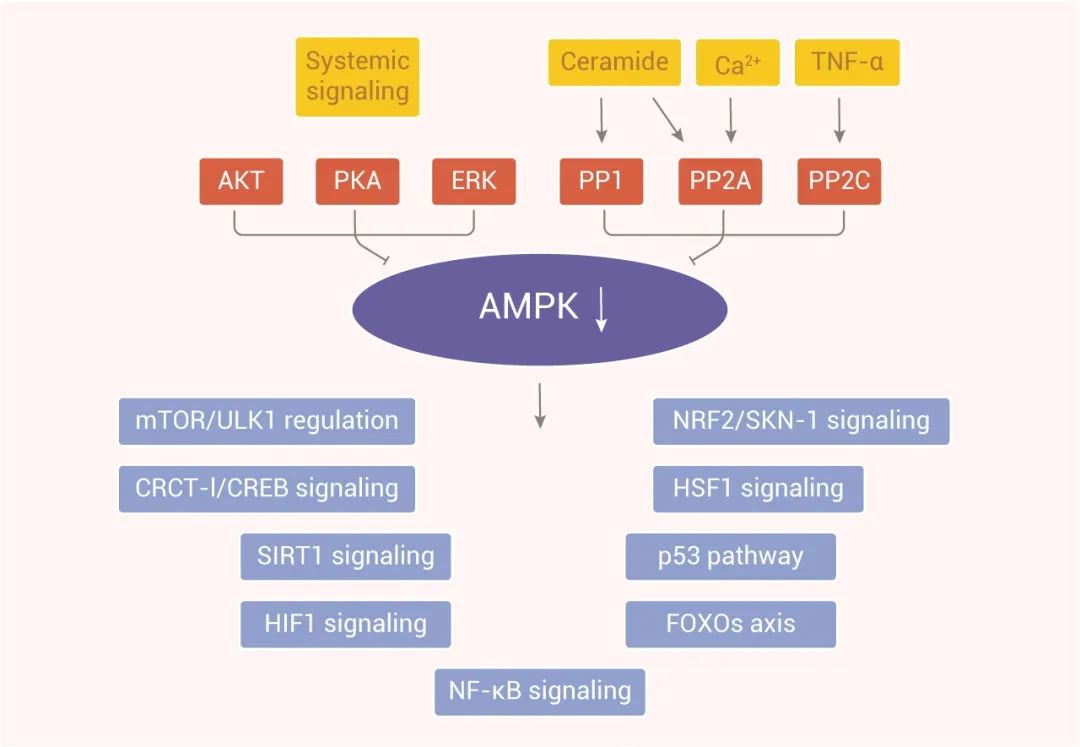
图 3. AMPK 激活的上游和下游信号通路的示意图[9]
总之,AMPK 的激活与许多延长寿命的途径有关,如抑制炎症,抑制 IIS、mTOR 信号,刺激 Sirtuin 信号,防止线粒体紊乱等。而 AMPK 激活剂 (如Metformin) 在减缓人类衰老过程中发挥了重要作用。
■ NF-κB 通路
NF-κB 系统是古老的宿主防御系统,涉及免疫应答,和多种外部和内部危险信号的应对,例如氧化应激,缺氧和遗传毒性应激等。许多研究表明,哺乳动物的衰老与 NF-κB 转录因子系统的激活有关。 免疫衰老是一种典型的与年龄有关的免疫系统功能下降,而 NF-κB 系统在调节先天免疫和适应性免疫方面均具有重要功能,例如 NF-κB 信号传导参与 T 细胞发育、活化、和增殖。
另外,NF-κB 还与 SASP 的控制有关。有意思的是,一些与长寿相关的基因能够抑制 NF-κB 信号传导,例如 SIRT1、SIRT6 和 FOXOs,它们通过抑制 NF-κB 信号延缓衰老过程,延长寿命。 由于 NF-κB 信号通路与信号网络有许多相互作用,因此抑制整个 NF-κB 系统可能是有害的。因此,开发针对 NF-κB 信号级联反应不同分支的特异性抑制剂至关重要。 ■ Sirtuins 通路
Sirtuins 在调节多种细胞过程中非常重要,如代谢、线粒体内稳态、自噬、DNA 修复、凋亡、氧化/抗氧化平衡和衰老。其中,SIRT1 催化组蛋白 H1、H3 和 H4 以及非组蛋白脱乙酰化,包括 p53、Ku70、FoxOs、PGC1-a、PPAR-γ 和 NF-κB,并通过这些底物的乙酰化和脱乙酰化改变它们的转录和酶活性以及蛋白质水平,广泛参与调节细胞衰老和生物体寿命。除此之外,SIRT1 还可通过整合多种信号参与衰老调节。 SIRT1 负向调节 NF-κB 信号。上文提到 NF-κB 信号在许多与炎症和衰老相关的疾病中被激活,并促进衰老过程。研究表明,SRT1720、白藜芦醇均可通过激活 SIRT1,抑制 NF-κB 的活性,在抗炎和衰老过程中起着重要作用。 另外,SIRT1 与 AMPK 在调节能量、新陈代谢和衰老方面有密切的相互作用,它们可以相互增强对方的活性,二者“互帮互助”。如 AMPK 通过诱导 NAD+/SIRT1 和上调自噬来延缓细胞衰老。SIRT1 还和 mTOR 通过调节自噬功能参与衰老调节,如 SIRT1 通过阻断 mTOR 通路恢复了氧化应激诱导的自噬损伤,提高了胚胎干细胞的存活率,有研究表明,白藜芦醇可抑制 mTOR 活性并改善细胞衰老。
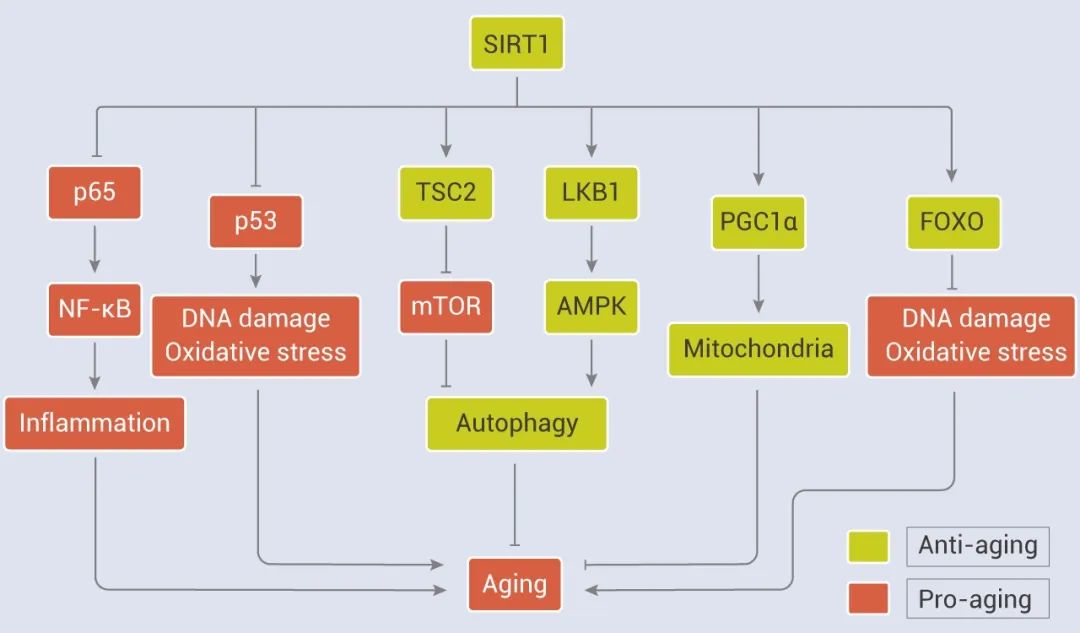
总之,SIRT1 信号通路与 NF-κB、FOXOs、p53、AMPK、mTOR、PGC1α 等许多重要通路密切相关,这些通路都与衰老及衰老相关疾病的发生有关。SIRT1 的上调可以影响衰老和衰老相关疾病中这些途径的调节。因此,用小分子激动剂激活 SIRT1 有望成为延长寿命、改善衰老相关疾病的有效手段。 另外,Sirtuin 家族成员 SIRT6 也被认为是衰老和衰老相关疾病的关键因素,但它在细胞衰老中的作用尚未清楚。
总结:衰老相关信号途径的确定扩展了可能刺激寿命途径或抑制衰老途径的小分子潜在靶标。影响衰老的化合物可以帮助预防多种与年龄有关的疾病,包括癌症和神经退行性疾病。相信随着对与衰老相关的信号网络日益深入的探索,改善老年健康,改变老年医学时日可期。
| 抗衰老化合物 |
| Picropodophyllin 选择性的胰岛素样生长因子-1 受体 (IGF-1R) 抑制剂,IC50 为 1 nM。 |
| Rapamycin 有效且特异性的 mTOR 抑制剂;与 FKBP12 结合且抑制 mTORC1;自噬 (autophagy) 激活剂;免疫抑制剂。 |
| Metformin 抑制肝脏中的线粒体呼吸链,导致 AMPK 活化,增强胰岛素敏感性,可用于 2 型糖尿病的研究。 |
| Sodium Salicylate NF-κB 抑制剂,可降低炎症基因的表达,促进老化肌肉的修复。 |
| BAY11-7082 IκBα 磷酸化和 NF-κB 抑制剂,选择性且不可逆地抑制 TNF-α 诱导的 IκB-α 磷酸化,并减少 NF-κB 和粘附分子的表达。 |
| Resveratrol 天然多酚,具有抗氧化、抗炎、保护心脏和抗癌的特性;它的靶点广泛,如 mTOR、JAK、β-amyloid、Adenylyl cyclase、IKKβ、DNA polymerase;特异性的 SIRT1 活化剂;Nrf2 激活剂,在小鼠模型中可以改善衰老相关的进行性肾损伤。 |
| 抗衰老化合物库 (Anti-Aging Compound Library) 收录了 2500+ 种化合物,主要靶向 Sirtuin、mTOR、IGF-1R、AMPK、p53、Telomerase、Mitophagy、Mitochondrial Metabolism、COX、Cytochrome P450、Oxidase 等靶点,是研究抗衰老机制的重要工具。 | CCK8 Cell Counting Kit-8,简称CCK-8试剂盒或CCK8试剂盒,是一种基于WST-8而广泛应用于细胞活性和细胞毒性检测的快速、高灵敏度试剂盒。 | |
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务
参考文献
↓ 下滑查看更多文献
1. Covarrubias AJ, et al. Senescent cells promote tissue NAD+ decline during ageing via the activation of CD38+ macrophages [published correction appears in Nat Metab. 2020 Dec 10;:]. Nat Metab. 2020; 2(11): 1265-1283. 2. Chini CCS, et al. CD38 ecto-enzyme in immune cells is induced during aging and regulates NAD+ and NMN levels. Nat Metab. 2020; 2(11): 1284-1304.
3. Slack C, et al. Signal Transduction Pathways in Ageing. Subcell Biochem. 2018; 90: 323-350. 4. Ai-Jun Ding, et al. Current Perspective in the Discovery of Anti-aging Agents from Natural Products. Nat Prod Bioprospect. 2017 Oct; 7(5): 335-404. 5. Zhang X, et al. Akt, FoxO and regulation of apoptosis. Biochim Biophys Acta. 2011; 1813(11): 1978-1986.
6. Weichhart T. mTOR as Regulator of Lifespan, Aging, and Cellular Senescence: A Mini-Review. Gerontology. 2018; 64(2): 127-134.
7. Laberge RM, et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol. 2015; 17(8): 1049-1061. 8. Herranz N, et al. mTOR regulates MAPKAPK2 translation to control the senescence-associated secretoryphenotype [published correction appears in Nat Cell Biol. 2015Oct;17(10):1370]. Nat Cell Biol. 2015;17(9):1205-1217. 9. Salminen A, et al. Age-related changesin AMPK activation: Role for AMPK phosphatases and inhibitory phosphorylationby upstream signaling pathways. Ageing Res Rev. 2016; 28: 15-26. 10. Salminen A, et alNF-kappaB signaling in the aging process. J Clin Immunol.2009;29(4):397-405. 11. Osorio FG, et al. NF-κB signalingas a driver of ageing. Int Rev Cell Mol Biol. 2016;326:133-174. 12. Chen C, et al. SIRT1 and aging relatedsignaling pathways. Mech Ageing Dev. 2020; 187: 111215.
13. Greer EL, et al. Signaling networks in aging. J Cell Sci. 2008; 121(Pt4): 407-412.





 电议 型 号:
电议 型 号:

