

企业档案
会员类型:会员

已获得易推广信誉 等级评定
(0 -40)基础信誉积累,可浏览访问
(41-90)良好信誉积累,可接洽商谈
(91+ )优质信誉积累,可持续信赖
易推广会员:8年
工商认证 【已认证】

最后认证时间:
注册号: 【已认证】
法人代表: 【已认证】
企业类型:生产商 【已认证】
注册资金:人民币万 【已认证】
产品数:91228
参观次数:11301684
公司新闻
高分文献:解析长寿与抗衰老背后的机制 - MedChemExpress
点击次数:299发布时间:2022/11/28
何为衰老
衰老取决于无法抵抗的时间推移,这是生物体自发的必然过程,表现为结构的退行性变性和生理功能的衰退。衰老会增加患癌症、糖尿病、心血管疾病等恶性或者慢性疾病的风险,并且也会降低对外界伤害的抵抗能力。例如,老年人由于免疫系统功能衰减,SARS-CoV-2 病毒感染后初始警报信号传递较慢,病毒容易在体内快速增殖 (图 1),COVID-19 感染在老年人中症状更严重[1]。了解衰老的机理不仅对于延长人类寿命有指导意义,还对老年人疾病的预防和治疗有重要的临床意义,也能提高老年人生活质量和幸福感。
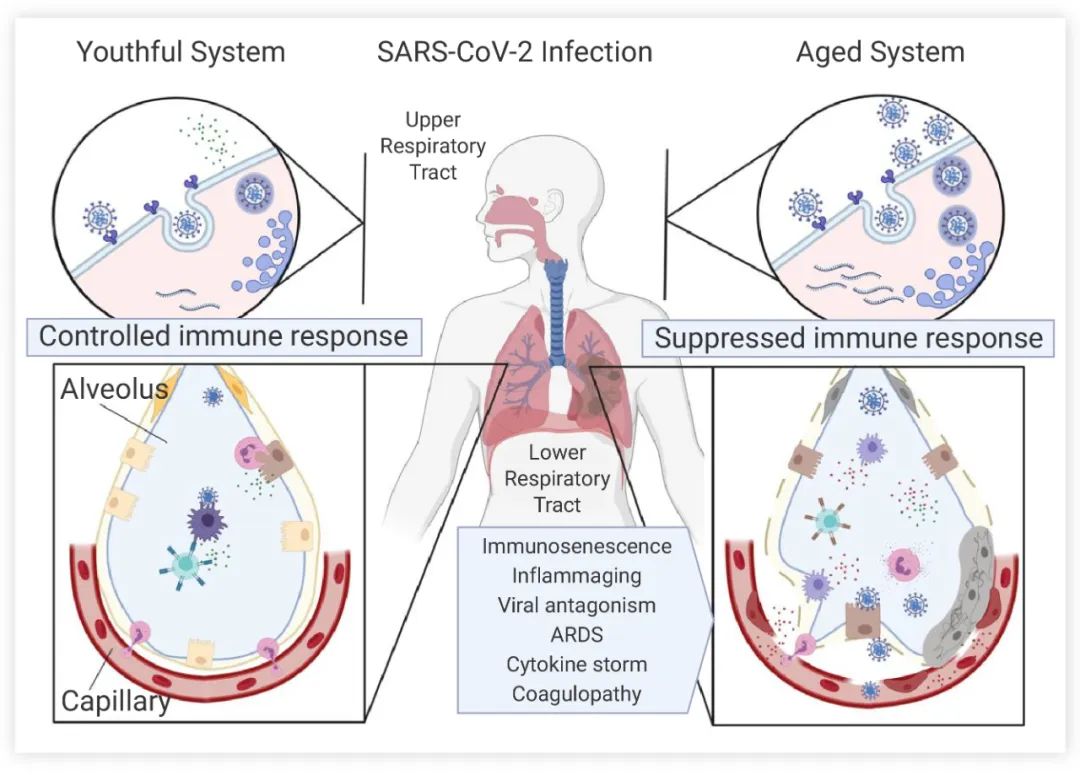
图 1. 年轻人与老年人的免疫系统对 SARS-CoV-2 的清除能力[1]
SARS-CoV-2 病毒在上呼吸道气道上皮细胞被内吞后,病毒传播到肺泡能被巨噬细胞和树突细胞识别。在老年的机体内,病毒警报信号*初较慢,病毒大量复制。有缺陷的巨噬细胞和受体数量有限的 T 细胞杀伤效果较差(右下),更多的细胞被感染,引发高水平的炎症细胞因子风暴。
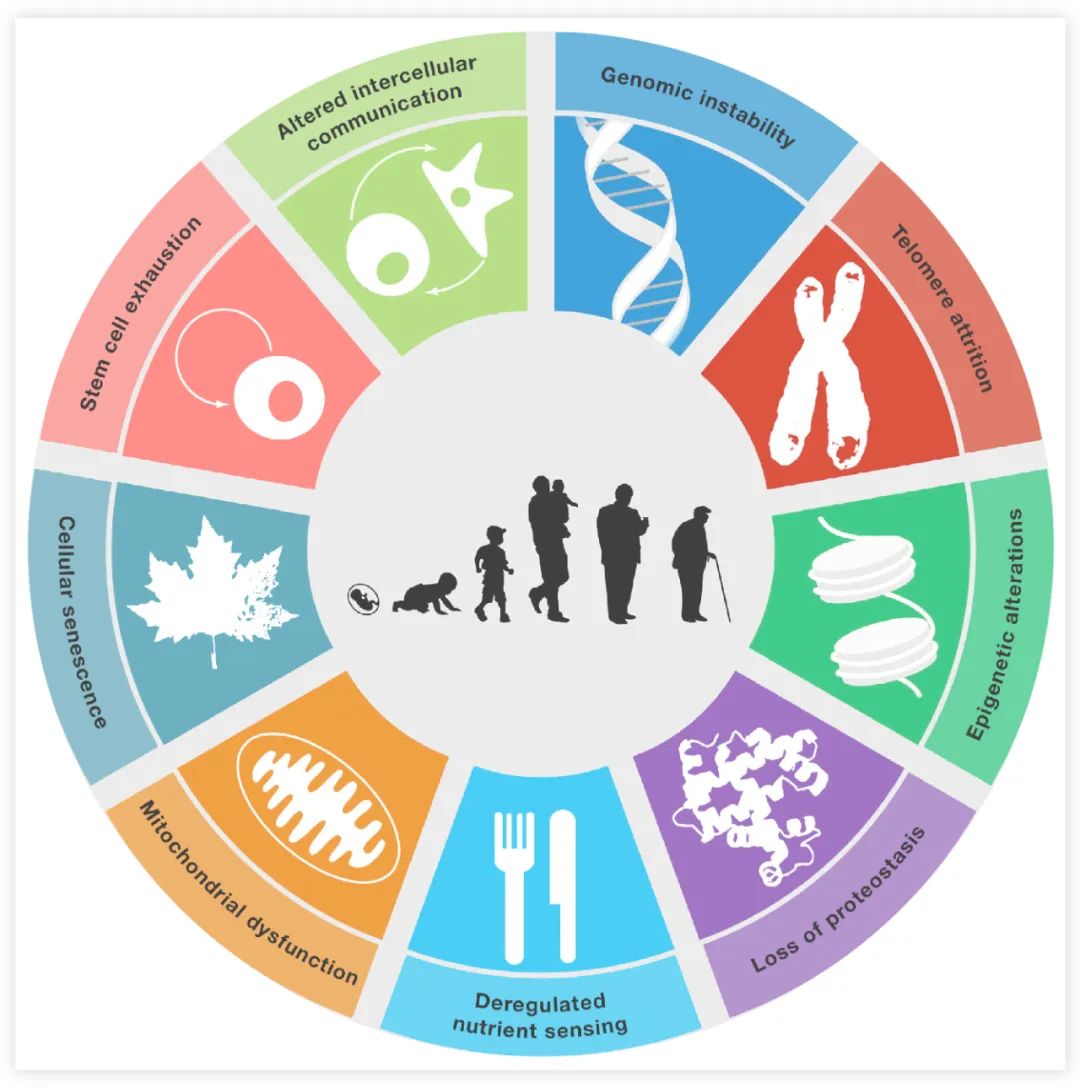
衰老是任何人都不可避免的,随着时间的推进,对于 “老” 的定义也在不断更新,衰老领域的研究也不会停滞不前,让我们来看看*近又有什么新发现~
研究“衰老”,探索长寿的秘密
-
长寿与低活性核糖体途径
核糖体生物合成和蛋白质合成是高度消耗能量的细胞过程,相关基因的低表达可减少合成并节省能量,同时也意味着一些诱发疾病的基因也不会过表达。在其他模式生物中也证明了,抑制核糖体通路能延长寿命。例如,使用 RNAi 抑制线粒体核糖体基因 mrps (mitochondrial ribosomal proteins) 延长了线虫的寿命[7]。
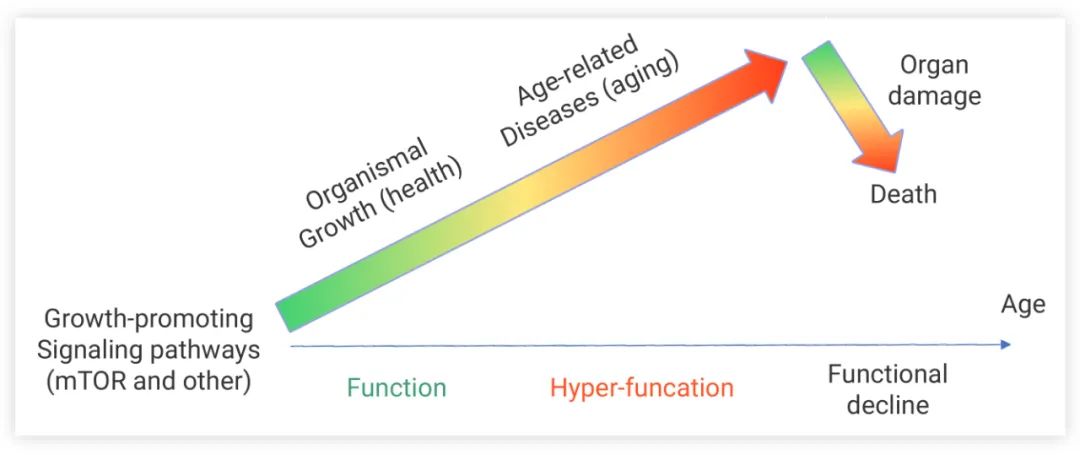
图 3. 细胞机能亢奋理论示意图[4]
衰老是由不必要的、持续激活的信号通路 (如 mTOR) 引起的。这些信号通路的激活直接导致与衰老有关的疾病发生与发展。
孔庆鹏研究员团队发现,与核糖体途径相关的基因,尤其是核糖体蛋白基因 (RPGs),在这些女性长寿老人中显著下调。下调 RPG 的启动子中,得分高的 TF 基序涉及 ETS 家族,ETS1 是其中表达差异*显著的基因。ETS1 基因与 RPGs 正向共表达 (positively co-express),其低表达能下调 RPGs。该基因被确定为是长寿老人中 RPG 转录的潜在关键转录因子。

图 4. 细胞水平下调 ETS1 的功能研究[6]
a. ETS1 敲低的细胞中衰老细胞 (β-半乳糖苷酶染色) 显著减少;b. 在 ETS1 敲低的细胞中,衰老调控因子 CDKN2A (p16) 和 CDKN1A (p21) 表达显著降低;c. 在 ETS1 敲低的细胞中,衰老相关分泌表型 (SASP) 基因表达显著降低
抗衰老与 BDNF-TrkB 通路
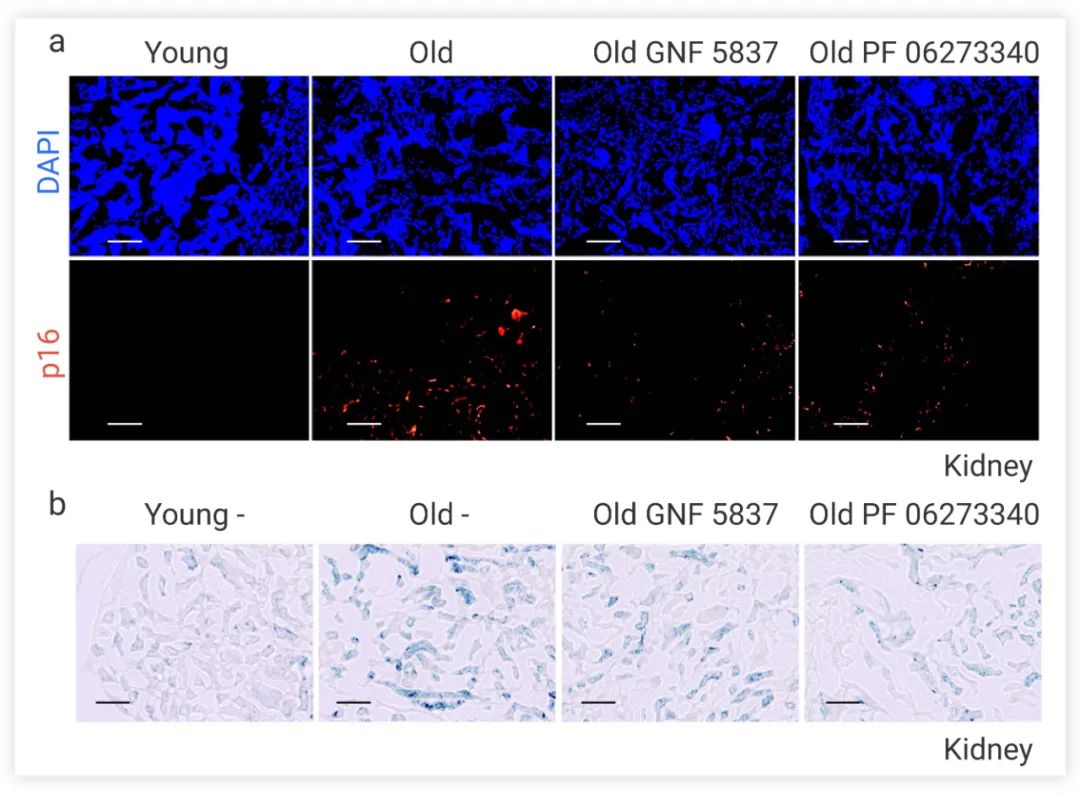
图 5. 注射 GNF 5837 和 PF 06273340 后自然衰老小鼠的肾脏中衰老标志物 p16 (a) β-半乳糖苷酶 (b) 水平显著降低[8]
衰老机制研究的“千层套路”
接下来是干货时间~
无论是研究衰老表型还是衰老背后的机制,通常会在体内外两个水平进行研究,研究者会使用模式动物 (表 1) 或细胞来模拟人的衰老过程和表型。

表 1. 衰老研究中常用的模式动物[9]
研究衰老,一步就是要对细胞或动物模型进行衰老诱导。在细胞中可使用各类抗生素 (如依托泊苷[8]),以及辐照 (诱导 DNA 损伤)、双氧水 (H2O2)[8]等。在动物中也可使用辐照[10]、抗生素 (如曲美替尼[10])、D-半乳糖[11]等。如要验证某基因对衰老的作用,可在细胞或动物模型中进行敲低或敲除,例如上文孔庆鹏研究员团队使用 siRNA 敲低 EST1 来验证该基因对衰老的影响[6]。

图 6. 细胞衰老时细胞内发生的变化[12]
包括DNA损伤、细胞增殖停止、应激反应蛋白的表达 (p16、Rb、p53 和 p21)、pH 为 6 时溶酶体膨胀 β-半乳糖苷酶染色为阳性;SASP 的产生和分泌增加
| 相关产品 |
3,400+ 种化合物,主要靶向 Sirtuin、mTOR、IGF-1R、 AMPK、p53、Telomerase、Mitophagy、 Mitochondrial Metabolism、COX、Cytochrome P450、Oxidase 等靶点,是研究抗衰老机制的重要工具。 |
| |
| |
| |
| |
参考文献
下滑查看更多↓
相关产品
-
 电议 型 号:
电议 型 号: -
电议 型 号:
-
电议 型 号:
-
电议 型 号:
-
电议 型 号:


