公司新闻
胞葬作用 (Efferocytosis) :程序性死亡细胞临终后一站 - MedChemExpress
点击次数:359发布时间:2022/11/30
人体每天都会产生上亿个死亡细胞。将旧细胞清除,给新的细胞留出位置,才能够使组织或器官保持活力与稳态。如果清除过程不顺利,则有可能会导致不良后果,如系统性红斑狼疮诱因就是凋亡细胞的积累诱发细胞继发性坏死,释放与损伤相关的分子模式,导致大范围的炎症反应。因此,清除死亡细胞对维持体内平衡具有重要意义。
胞葬作用 (Efferocytosis) 就是指吞噬细胞清除程序性死亡细胞的过程。在大多数组织中,胞葬作用由专职性吞噬细胞 (巨噬细胞和树突细胞) 或垂死细胞附近的非专职性吞噬细胞 (上皮细胞和纤维细胞) 来完成。胞葬作用能够阻止垂死细胞发生继发性坏死,以免释放出可能引起炎症的有害细胞内容物 (如氧化物和蛋白酶)。那么吞噬细胞是如何识别垂死细胞并清除掉它们的呢? 来听听胞葬作用的三部曲~Find-Eat-Digest~♪
垂死细胞会释放出可溶性介质,这些介质中就包含让吞噬细胞识别的 “Find me” 信号,包括核苷酸 (如 ATP, UTP)、膜脂 (如磷酸鞘氨醇)、趋化因子 (如 CX3CL1) 等。这些信号不仅能吸引吞噬细胞,还能使吞噬细胞做好 “作战” 的准备,例如增强吞噬受体和消化机制的表达。
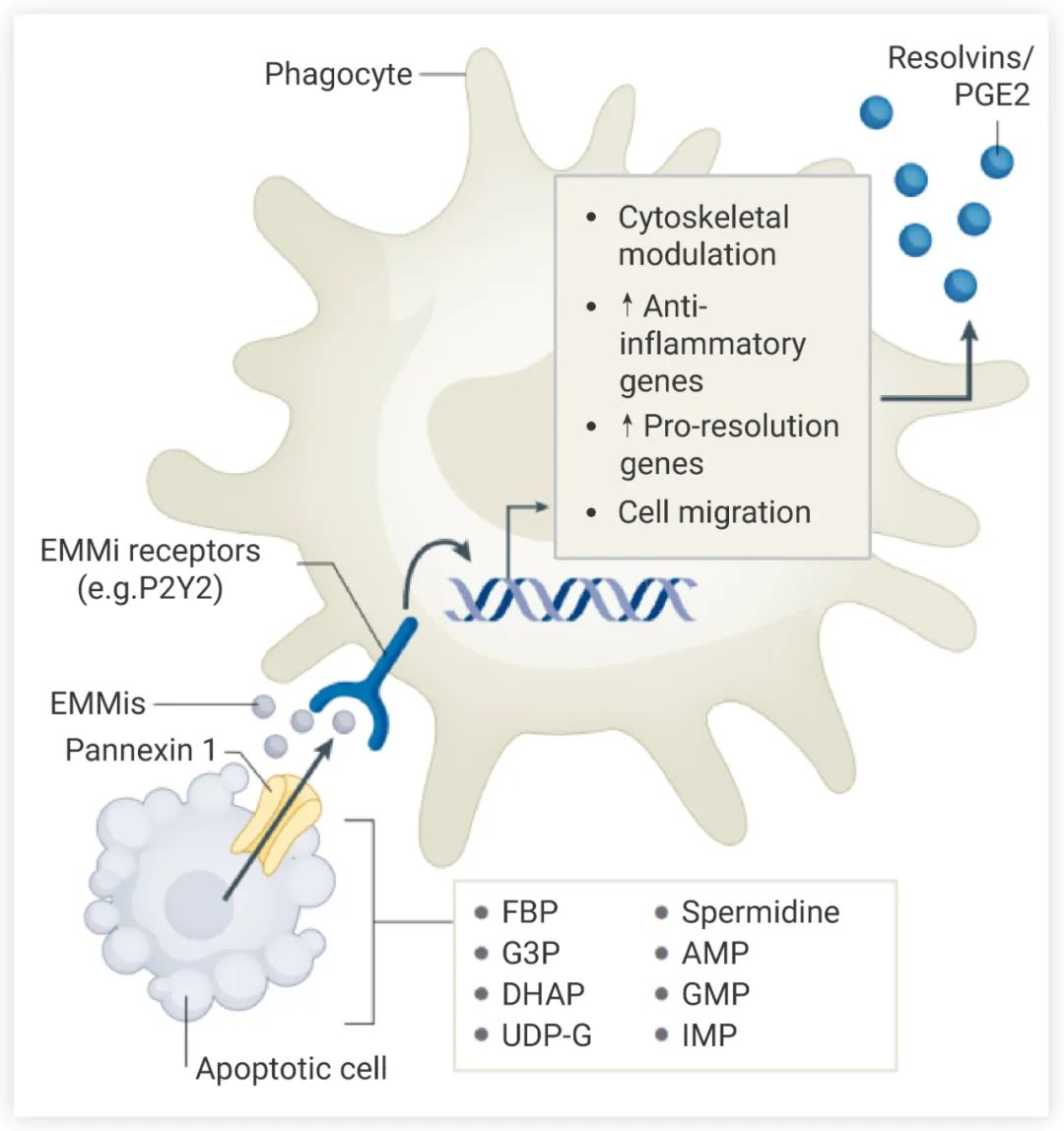
凋亡细胞的核苷酸 “Find me” 信号可通过 pannexin 通道 (被凋亡过程中 caspase 3/7的裂解激活) 释放 (图 1)。释放的 ATP 通过嘌呤受体 P2Y 诱导吞噬细胞迁移。此外,核苷酸还能够调节巨噬细胞的免疫机制。例如,凋亡细胞的 ATP 或 AMP 可转化为腺苷,腺苷再通过腺苷受体抑制炎症,上调抗炎 (anti-inflammatory) 和促消炎 (pro-resolution) 基因表达 (包括 Nr4a1 和血小板反应蛋白)。非凋亡细胞的脂膜完整性被破坏,炎症信号直接被释放到细胞外。被病原体感染的细胞会释放病原体相关分子模式 (PAMP),该信号可与吞噬细胞上或吞噬细胞中的模式识别受体 (PRR) 结合,影响巨噬细胞的功能和免疫激活。另外,非凋亡细胞也会释放损伤相关分子模式 (DAMP),触发炎症反应,可作为巨噬细胞的趋化因子。图 1. 吞噬细胞识别凋亡细胞[1]
凋亡细胞释放 “Find me” 信号 (核苷酸、膜脂、趋化因子等),吞噬细胞上相应的受体与之结合,诱导吞噬细胞迁移、上调抗炎/促消炎基因表达,以及调节细胞骨架吞噬细胞迁移到凋亡细胞附近后,表面受体识别凋亡细胞的 “Eat me” 信号,与之结合。“Eat me” 信号包括磷脂酰丝氨酸 (Phosphatidylserine, PS)、钙网蛋白、氧化低密度脂蛋白等,其中 PS 是*有效、进化上*保守的信号分子。所有形式的细胞死亡的一个共同特征是质膜中磷脂不对称性的丧失,PS就会暴露在细胞外,促进死亡细胞被吞噬。凋亡过程中,位于细胞内侧的 PS 会在早期阶段从内侧翻转到表面。吞噬细胞的 PS 受体可以直接或间接识别暴露在表面的 PS。BAI1 (PS 受体) 与 PS 结合后,通过 ELMO1-DOCK 复合物启动细胞内信号传导,以诱导 Rac1 介导的肌动蛋白细胞骨架重排 (图 2),为 “吃掉” 凋亡细胞做好准备。
与之相对的是,健康细胞表面暴露 “Don't eat me” 信号,包括 CD47 和 CD24。活细胞上的 CD47 与巨噬细胞的 SIRPα 结合,导致 SIRPα 胞质结构域的酪氨酸磷酸化,募集到的 SHP1/2 再通过非肌肉肌球蛋白 IIA 来抑制吞噬作用 (图 2)。这些信号能使健康细胞避免被吞噬细胞清除。
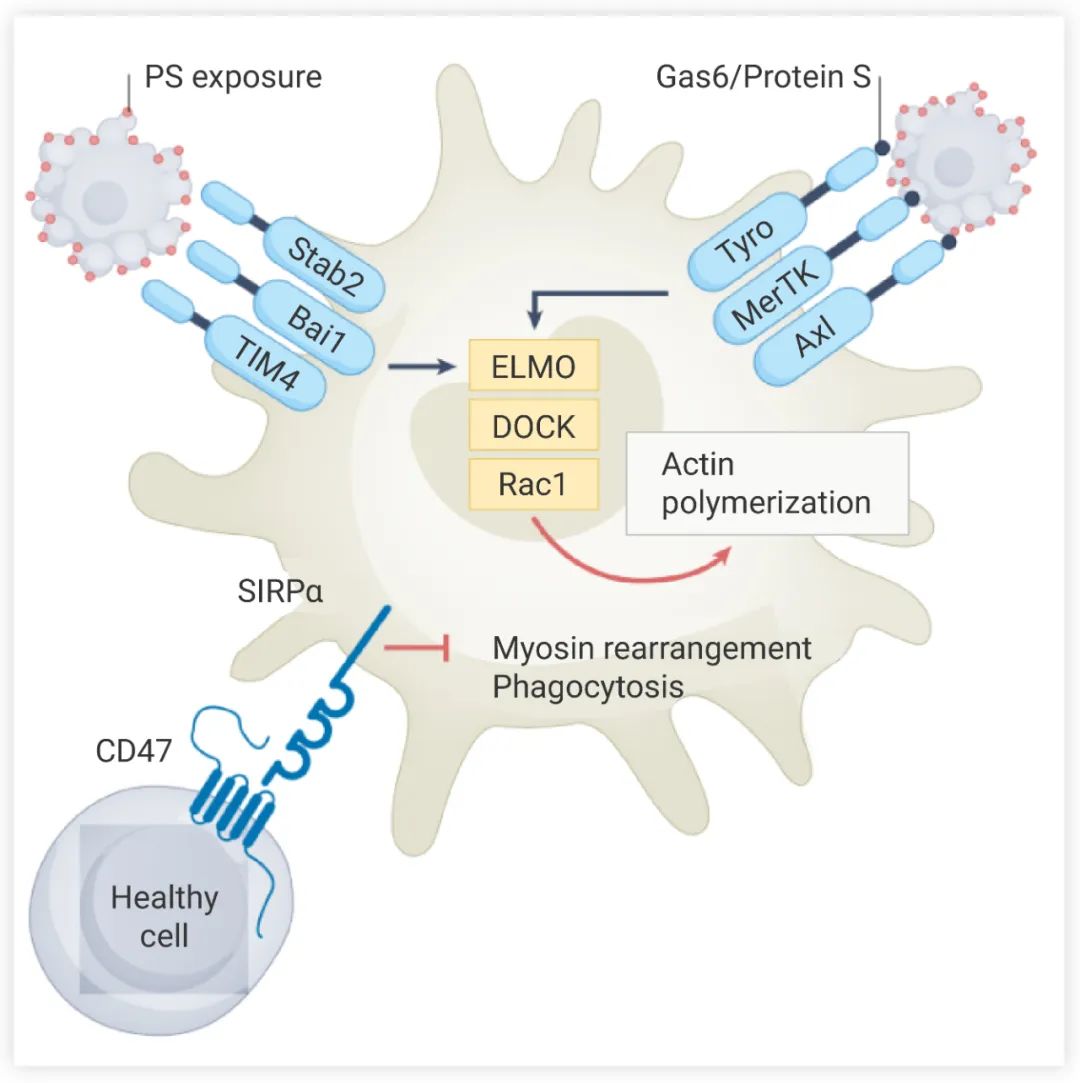
图 2. 吞噬细胞结合凋亡细胞[1]
吞噬细胞直接 (BAI1, TIM1) 或间接 (AXL, MerTK) 识别凋亡细胞表面的 PS,激活 ELMO 和 DOC 蛋白的表达,进而激活 GTPase RAC1,增强肌动蛋白重塑和吞噬杯的形成。健康细胞表面的 CD47 能阻止被吞噬
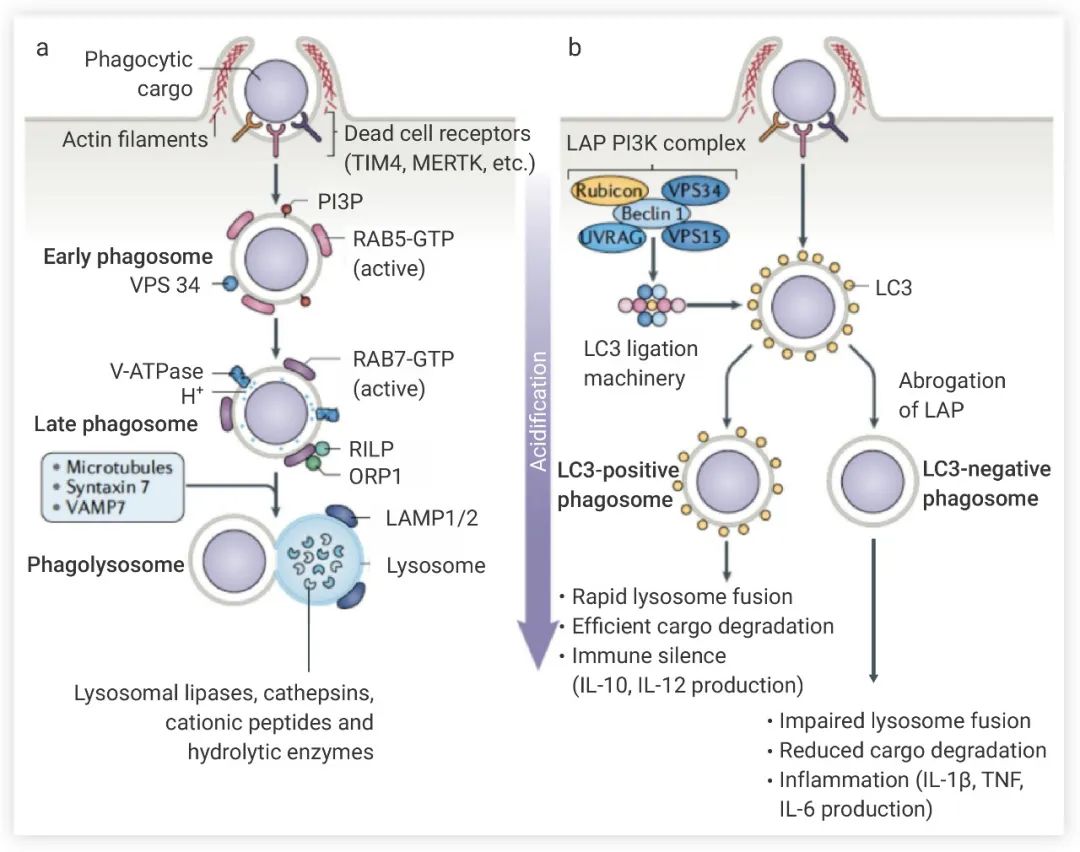
在与垂死细胞结合后,吞噬细胞启动肌动蛋白重塑,质膜内陷及局部外溢形成吞噬体,通过胞吞作用将垂死细胞 “吃掉”。胞吞后形成吞噬体,吞噬体膜会发生多种生化变化,这些过程由 Rab GTPase 蛋白家族控制。吞噬体成熟后,通过形成 Ca2+ 依赖性 SNARE 复合物 (由 VAMP7 和 Syntaxin 7 组成) 与溶酶体直接融合 (图 3a)。溶酶体内有大量蛋白酶、核酸酶和脂肪酶,可消化吞噬体中凋亡细胞。另外还有 LC3 (微管相关蛋白 1A/1B-轻链 3) 相关吞噬作用 (LAP) 通路 (图 3b)。吞噬细胞吞噬垂死细胞后,LAP PI3K 复合物被募集到含有凋亡细胞的LAP相关吞噬体 (LAPosome),该复合物对 LAPosome 的保持至关重要,并且激活了 LC3 的连接机制。LC3 的连接促进了 LAPosome 与溶酶体融合,提高了清除死细胞的效率并且保持免疫沉默。图 3. 吞噬细胞吞噬凋亡细胞[2]
a. 溶酶体降解途径:VPS34 被募集到RAB5-positive早期吞噬体,催化磷脂酰肌醇形成 PI3P (这些都是吞噬体成熟所必需的物质),早期吞噬体成熟为晚期吞噬体后,*终与溶酶体融合。b. LAP 通路:LAP PI3K 复合物 (由 Beclin 1、VPS34、VPS15、UVRAG 和 Rubicon 组成) 激活 LC3 与 LAPosome 的连接,促进 LAPosome 的成熟,以及与溶酶体融合
溶酶体中含有大量蛋白酶、核酸酶、脂肪酶。吞噬体与溶酶体结合后,形成新的吞噬溶酶体 (Phagolysosome)。高酸度的环境 (pH=4.5~5.0) 和活性组织蛋白酶能降解垂死细胞。吞噬细胞还可重利用一些降解的垂死细胞成分。大部分情况下,死亡细胞的数量都是远远超过吞噬细胞,所以吞噬细胞需要同时清除多个细胞。而垂死细胞带有膜、胆固醇、蛋白质、核酸等,这些物质需要被代谢。这对于吞噬细胞来说是巨大的负担,需要控制其自身的体积和表面积以及迅速改变免疫代谢环境。凋亡细胞本身也能改变吞噬细胞的免疫代谢 (图 4):(1) 凋亡细胞表面的 PS 与受体结合后能激活或上调 LXR 和 ABCA1,可促进胆固醇的代谢与输出;(2) 凋亡细胞线粒体的脂肪酸氧化可促进抗炎因子 IL-10 的表达;(3) 吞噬细胞利用凋亡细胞的精氨酸 (Arginine) 驱动肌动蛋白细胞骨架重排,促进之后几轮的凋亡细胞摄取;(4) 糖酵解与葡萄糖输入的增加也促进 ATP 生成和肌动蛋白聚合。
图 4. 凋亡细胞调节吞噬细胞免疫代谢环境[1]
常规的胞葬作用研究是在体外进行,例如使用原代人巨噬细胞和凋亡 Jurkat T 细胞进行免疫荧光染色后,可在显微镜下观察到胞葬作用的过程[3]。但由于凋亡细胞易被清除,在体内检测仍是个巨大的挑战。不过,办法总比困难多嘛~
Raymond 等人开发了一种基因编码的荧光报告基因 CharON,实现了在果蝇胚胎发育过程中胞葬作用的活细胞追踪[4]。这一研究结果于今年 3 月发表在 Science 杂志上。
研究者们首先设计了针对细胞凋亡和胞葬作用的传感器。针对细胞凋亡,他们设计了 一种绿色荧光蛋白 (GFP) 探针。细胞凋亡时表达的 caspases 3/7 能切割 GFP,使之发出荧光 (图 5a);GFP 在溶酶体的酸性条件下易淬灭,经突变 Q204H 基因后, GFP 对 pH 耐受。经过以上改造,他们设计出 pH 耐受的绿色荧光探针 pH-CaspGFP——它能准确地报告凋亡的细胞。针对吞噬作用,他们设计了一种新的红色荧光 pH 值传感器 pHlorina,它随着 pH 值的降低而增加荧光强度。*后将这两种传感器结合起来,获得探针 “CharON” (图 5b)。CharON 转基因的细胞,在凋亡时首先发绿色荧光,在被吞噬并在溶酶体内酸化后,随着 GFP 逐渐淬灭,pHlorina 的红色荧光强度增加。CharON 将连续性细胞事件可视化。
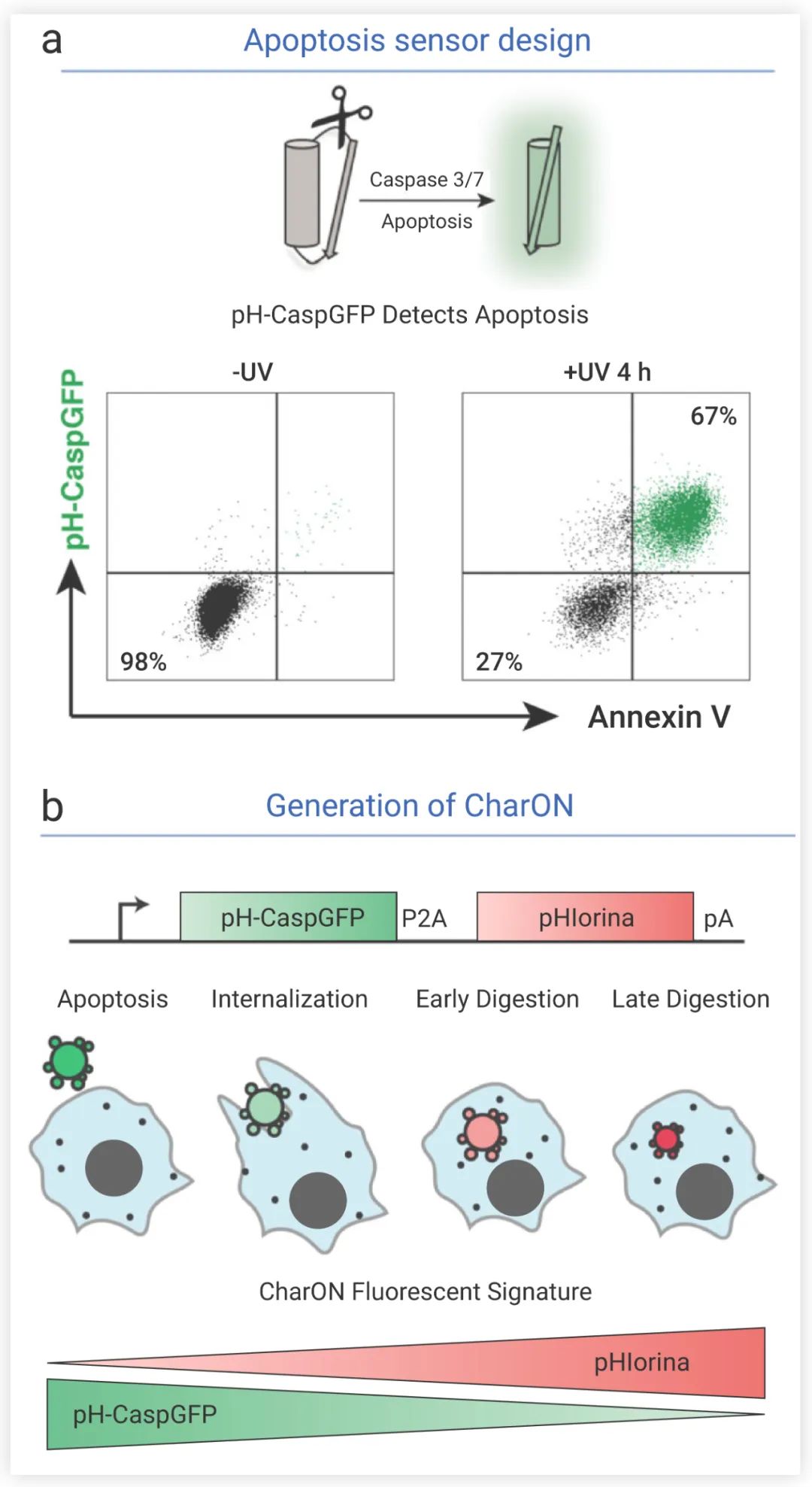
图 5. pH-CaspGFP 的设计 (a) CharON 探针检测胞葬过程 (b) 示意图[4]细胞凋亡时,caspase 3/7 切割 GFP,发绿色荧光;被吞噬细胞吞噬后,在酸性环境中 GFP 逐渐淬灭,红色荧光 pHlorina 强度增加为了追踪体内胞葬过程,研究者培育出 CharON 转基因果蝇。在果蝇胚胎发育的中后期,发育中的中枢神经系统 (CNS) 有一波细胞凋亡。CharON 使体内胞葬过程更完整地呈现出来,包括细胞凋亡、吞噬细胞募集、Find、Eat 和 Digest (图6)。通过 CharON 还观察到,胚胎果蝇 CNS 中清除凋亡细胞的“重任”由具有吞噬功能的神经胶质细胞和分散的腹侧血细胞 (巨噬细胞) 共同承担。并且还观察到神经胶质细胞和巨噬细胞中吞噬体的大小也有差异,巨噬细胞中凋亡细胞的荧光更密集。
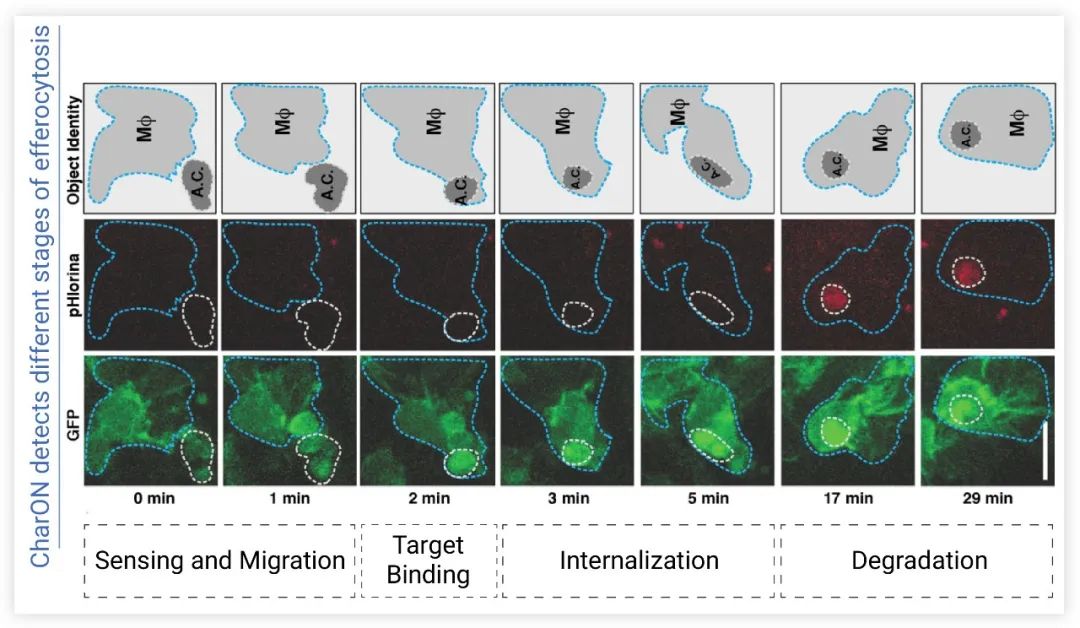
图 6. CharON 可视化果蝇胚胎内胞葬作用的不同阶段[4]
0~1 min:凋亡细胞募集巨噬细胞;2 min:凋亡细胞与巨噬细胞结合;3~5 min:巨噬细胞吞噬。MΦ (蓝色虚线轮廓):GFP 标记的巨噬细胞;A.C. (虚线轮廓):凋亡细胞。细胞凋亡诱导 pH-CaspGFP (绿色) 激活和巨噬细胞 (绿色) 迁移,靶标与受体结合后被吞噬,消化后通过增加 pHlorina 信号 (红色) 检测凋亡细胞的酸化/降解胞葬作用涉及多个信号通路,包括受体网络、细胞溶质信号分子、细胞骨架快速重排、凋亡细胞消化和免疫代谢通路。这些通路运行不畅则有可能导致多种胞葬缺陷相关疾病 (神经退行性疾病、视网膜变性、动脉粥样硬化、癌症等),因此了解胞葬作用机制对于治疗这些疾病更有重要意义。尽管研究之路还很漫长,但相信总有一天我们能攻破难题~
| 相关产品 |
| 可方便、快捷的检测细胞凋亡。经 Annexin V-mCherry 染色后,正常细胞基本无荧光,凋亡细胞及坏死细胞呈红色荧光。 |
| 细胞凋亡与坏死检测试剂盒 采用 Hoechst 33342 和碘化丙啶(Propidium Iodide, PI)双染的方法快速检测细胞凋亡与细胞坏死。 |
| DMUP 是一种有效的 CD47-SIRPα axis 抑制剂。DMUP 在 A549 细胞中诱导细胞凋亡 (apoptosis) 并增加巨噬细胞的吞噬作用。 |
| Neriifolin 可靶向 Beclin 1,抑制 LC3 相关吞噬体的形成,改善实验性自身免疫性脑脊髓炎 (EAE) 的发展。 |
| Pennogenin 3-O-beta-chacotrioside 是从七叶一枝花中分离得到的活性物质,可以调节自噬 (autophagy),增加自噬相关蛋白 LC3 和 Beclin-1 的表达。 |
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务
下滑查看更多↓
1. Parul Mehrotra, Kodi S. Ravichandran. Drugging the efferocytosis process: concepts and opportunities. Nat Rev Drug Discov. 2022; 21(8): 601-620.2. Emilio Boada-Romero, et al. Mechanisms and physiology of the clearance of dead cells by efferocytosis. Nat Rev Mol Cell Biol. 2020 Jul; 21(7): 398-414.3. Evans AL, Heit B, et al. Quantitative Efferocytosis Assays. Methods Mol Biol. 2017;1519:25-41.4. Raymond MH, Ravichandran KS, et al. Live cell tracking of macrophage efferocytosis during Drosophila embryo development in vivo. Science. 2022 Mar 11;375(6585):1182-1187.
相关产品






 电议 型 号:
电议 型 号:

